在基因水平上研究嗜中性白血球的基因表達變化提示精神修煉在多方面調控基因表達(圖)


李全貞1 李平2 加波瑞艾娜•加西亞2 理查德•強森2 封莉莉2
1: DNA微排列中心,免疫學中心,西南德克薩斯大學醫療中心
達拉斯,德克薩斯州75390
2: 貝勒醫學院,醫學系
休斯頓,德克薩斯州77030
摘要
在基因水平上,人類和其它物種有著令人不可思議的相似性,這促使我們另闢蹊徑去尋找與人的特性有關的,在基因調控中起重要作用的因素。越來越明顯的安慰劑效應說明人的思想(精神)能顯著影響其對於健康、疾病、和醫藥的反應。精神修煉和信仰在個體對於健康問題和疾病的反應中起著重要的作用,並將影響到個體的生活和幸福。因此,我們對一群進行精神修煉的人,在基因水平上研究了他們的基因表達狀況。和正常健康人相比,我們發現法輪功學員的嗜中性白血球的功能以及基因表達有著非常顯著的變化。這些變化表現為:吞噬和殺傷細菌的功能增強,和免疫力有關的基因表達增加,細胞死亡基因被雙向調控從而使得正常細胞存活期延長的同時又加速受刺激的細胞凋零死亡以便炎症迅速消除,以及細胞新陳代謝的下降等等。細胞新陳代謝的下降表現在泛素依賴性的蛋白質降解系統的相繼下調控,這種下調控可能是和細胞內的代謝產物減少相關,而非直接調控。細胞死亡的雙向調控可能是陰陽調和的表現,因為炎症狀態下的細胞死亡加速可能使機體在免疫力增強時沒有副作用-宿主的自傷。我們這裏提供了第一手證據,來證明精神修煉可能通過精神層面上對基因組的調節來改變免疫力、細胞死亡和新陳代謝率等等。這些發現說明了人的精神對機體的至關重要的調節作用,意味著我們急需建立更多的模型來研究意識、認知、和精神等人類特有的特性和行為等因素對基因調控的影響。
有多種因子在人類健康中起著重要作用。這些因子可能在心理上起作用從而影響免疫和神經內分泌系統[1,2]。2000年6月27、28日,在美國國立健康研究所(NIH)的Natcher會議中心召開了由該所主辦的題為「朝向高層分析」的研討會。會議的主題就是如何加強研究社會和文化對健康的影響。為使大家更好地認識到在健康調研中社會、行為和生物層面上分析研究的相互依賴性,會議強調了社會文化因素對健康和疾病的影響。會上,社會學家提出了作為單元分析的社會和文化體系,認為應該在包括宗教和精神活動等社會和人際關係如何影響健康這一方向上,進行更為廣泛的研究 [3]。長期以來,人們一直在猜測人的思想會顯著調控人體。不過,人的思想如何在分子和細胞水平上具體地調控基因表達,仍是我們需要研究的課題。
人類健康的關鍵因素─人性:都在於精神意識中
科學家在25年前報導說,不同物種間的差異可能只是源於基因表達數量的差異,而非基因的一級結構不同[4]。所有哺乳動物的基因庫是如此的相似以至於很難理解為何相似的基因物質會產生如此眾多的不同生物[5]。黑猩猩和人類就具有98.7%的相同基因物質,當然在形態和認知能力上,二者具有極為顯著的差異 [6]。Enand等人推測基因表達方式的差異性可能造成了物種之間的差異。他們發現大猩猩和人只在腦部的某些基因轉錄方式上有差異,且人腦的某些基因活性增強[6]。有些科學家認為應該用一個「轉(人類特異性)基因小鼠模型」來進一步研究是否某些人類特異性基因能把小鼠變得更像人。當然,也有的科學家覺得這樣的「魔術」基因根本就不存在。
那麼超越一維基因庫以外的「魔術」因素是甚麼呢?如果不同動物的基因如此類似,也許人與其他物種的差異性在於人的基因受高級思維調控的變化造成的,而這種變化發生的時間,部位,和強度可能會起著決定性的作用。人和大猩猩的大腦在基因活性上的差異說明了人類的特異性可能由人類特有的精神活動主宰,基因庫的一級結構不可能是唯一的因素。人類和其它生物的在疾病和健康上的巨大差異暗示我們肯定還有其它因素。對於物理和病理原因造成的基因調控,我們已有了相當多的了解,但是精神如何調控基因表達,哪些基因能受思想活動的影響,它們又是如何去進一不調控那些受物理和病理刺激調控的基因,對於這些問題我們卻是知之甚少。
「安慰劑/反安慰劑」效應的啟示
「安慰劑(Placebo, 源於拉丁文「我會高興」)效應」是指病人在使用無藥性片劑(如糖片)後產生的良好感覺。這種現象在醫學界隨處可見,以致很少有醫生對其提出挑戰[7-11]。而「反安慰劑(Nocebo,源於拉丁文「我會受傷」)效應」恰恰相反,它讓我們看到消極的想法會對「被試」產生不良影響 [12-15]。「安慰劑效應和反安慰劑效應」說明,病人的思想(精神)在其對於疾病和藥物治療的反應中會扮演重要的角色。
雖然很少有人對安慰劑效應提出異議[16],最近Petrovic等人明確提出,不管是安慰劑還是止痛片都能在大腦富有「鴉片感受體」的同一區域加強血液流動[17]。在隨後的一個帕金森氏綜合症的臨床病例和大多數抗抑鬱劑試驗中,發現安慰劑不僅緩解了病狀還改變了大腦的化學反應[18-20]。科研人員發現,與20年前相比,更多的病人在使用安慰劑後病情有好轉。這說明現代生活的緊張和環境的壓力可能引起了更多的與心理有關的疾病。Torrey和 Miller博士在他們的著作《看不見的瘟疫:從1750年至今精神病的增長》中總結到,「近幾十年來,至少在一些工業國家,確鑿的證據表明由沮喪和情緒低落引起的人類疾病有蔓延的趨勢,甚至是最嚴重的精神紊亂--精神分裂症」[21]。在一項針對2001年9月11日之後心理壓力反應的調查中, 「9/11後效應」進一步證實了這樣一種趨勢:即使遠離出事地點的人也表現有實質上的精神壓力症狀[22]。住在「雙塔」附近的應答者中,有20%的人表現出「重大創傷後遺症」(或稱:創傷後應激障礙,PTSD)23。在另一項針對988名曼哈頓居民的研究中發現,抽煙、喝酒和吸大麻的量都有顯著增加,而這可能與不同的心理狀況有關[24]。據教育局的研究表明,在一項針對110萬公立學校學生的調查中,約有75,000名兒童表現出六種以上的PTSD症狀,而這足以被診斷為精神紊亂[25,26]。
很明顯,安慰劑的治病效果是由於人的期望和信念能影響生理反應[27]。另外,安慰劑效應近年的增加也說明了現代社會人類的綜合疾病和心理壓力的增加可能與人類特有的心理因素有關。這就是說人類的疾病不僅僅源於簡單的病理、生理、物理刺激以及基因缺陷。因此,目前以動物模型為基礎的針對靶基因的研究就完全不應該成為研究人類特有疾病的病源學、病理學和心理學的唯一基礎。
在基因水平上測定與健康表型有關的基因表達狀況
所有的生物天然就應該有一個完整的,自我調控的健康的生活。不良的健康狀況不是一種自然狀態。一般說來,生物從本質上是不該生病的,如同呼吸、消化、分泌和生殖作用,生物體內就具備組織的自癒過程。健康是一種自然狀態,因為它是和諧的、平衡的。所以,確定在健康的人群中基因如何受心理和精神的調控非常重要,因為那些生物體系可能具有更高維的信息,從而保持健康和抗疾病能力增強。
基因表達的調控是生物學現象的一個重要方面。在對環境條件的反應,多細胞組織的發展,形態的變化,和病理的改變等方面雖然都有廣泛的研究,如何確定基因的病理學變化仍對研究人員是一個巨大的挑戰。在「疾病」的基礎上來確定發病有關的基因變化可能有不少問題,因為一些有利於機體的上調控或下調控可能被認為是有害的,反之亦然。動物模型是一個很好的研究系統。然而,與人性有關的複雜因素卻完全不能用動物來研究,因為人的認知、行為、和思維永遠不可能有動物模型,其中也包括對最高級的靈長類動物黑猩猩的研究。此外,目前尚無對於人的心理和精神特質的科學測量單位。
最常見的疾病反而最難研究。心臟病、癌症、糖尿病和精神病等等,所有這些都被認為是「複雜的」和「多因素」疾病,因為它們不是由單個基因的變異和單一外界條件所造成的。「候選基因逼近法」也受到批評,因為很多實驗結果沒有重複性,並且它無法包涵所有的成因基因和基因的多態現象[28]。那些綜合疾病實際上產生於很多基因的聯合作用,以及環境因素及冒險行為等[29-32]。在近期《科學》雜誌上,有多篇文章闡述了科研人員在研究綜合疾病時遇到的困難[29,33-35]。要想確定特定的基因導致了綜合疾病,就像大海撈針一樣難。因為即便能找到,這些基因往往只對這些複雜性疾病的形成起微小的作用。同時,這些作用的大小又受不相關基因和環境因素,以及人的思想(精神)的影響。經常是在一個試驗人群中確定的綜合疾病的易感基因座在別的人群中無法重複。越來越多的研究發現,對於那些綜合疾病,超過70%的中風、直腸癌、冠心病和第二型(成人型)糖尿病都可以通過改變生活方式而得到預防。這說明自我調控在預防此類綜合疾病中起著重要的作用[30,33]。看來,預防仍是最好的治療[36],因此我們亟需研究為甚麼有些人能保持健康,以及他們如何具有對疾病的抵抗力[37,38]。
人們對於身心緊密相關這一現象越來越感興趣。而早先,這一課題正是由於它的複雜性而被系統的科學研究排斥在外。身心醫學(MIND/BODY MEDICINE) 是建立在過去25年中2000多篇發表在由專家審定的雜誌上的科學研究的基礎上[27]。哈佛醫學院的HERBERT BENSON,這位「身心醫學」的先驅在這一領域中作出了巨大的貢獻。多年來積累下來的豐富證據表明(心靈)修煉所獲得的顯著的生理效應直接與思想及靈性有關。這類修煉包括宗教和非宗教修煉。在西方社會,宗教修煉佔主導地位,而東方文化則強調通過身體和精神的訓練來激活能量體系,比如瑜珈、氣功、太極以及最近出現的法輪功(FLG),又稱法輪大法[39-44]。正當瑜珈、打坐或祈禱的健身效果越來越多地被證實並逐漸被公眾所接受的時候。這些顯著效果在生理水平上的機理並未被人們廣泛認識,更不用說在細胞或分子水平上去證實和系統的研究。另一方面,生物研究技術的一些重大進步使我們可以對單個細胞內許多具體的變化進行快速和精確的檢測。這些科學技術的進展有利於研究這些修煉引起的身體在細胞或分子水平上變化。根據1999年初中國政府的一份調查,在中國大陸有7千萬到1億人煉法輪功。在所有旨在增進健康的不同方法中,法輪功在中國人中普及最快,影響力最大。1998年,在北京、武漢、大連地區和廣東省,由當地的醫療機構進行了五項獨立的健康調查。採樣人群超過35,000人,是迄今對法輪功修煉者最系統和全面的健康調查(https://www.pureinsight.org/sci/sci/eng/newscontent.asp?ID=14248)。為了探尋修煉人健康受益的分子機理,我們在基因水平上研究了法輪功修煉者嗜中性白血球的基因表達,主要的發現總結如下。
陰陽平衡基礎上的免疫調控
有證據表明心靈修煉和身心鍛煉方法能增進免疫系統的效力進而在臨床治療各種疾病中很有效 [45-47]。緊張和不良情緒會造成免疫紊亂,從而導致感染的加劇[2]。因此最好的防止傳染病流行的方法就是增加人群中具有天然抵抗力從而具有完全免疫力的人數。健康人具備天然免疫力不僅能使其具有完全免疫力,而且能減少人群中傳染病的流行。
最近,我們用DNA微排列技術比較了修煉人(n=4) 和非修煉 (n=4) 的健康人嗜中性白細胞的基因表達狀況,我們發現修煉人的嗜中性白細胞的基因表達的超常變化,此外還有明顯的功能上的變化。我們使用Affymnetrix 的載有12,000個探針的基因芯片發現了有眾多的基因在表達水平上有相當大的改變。其中有兩倍以上變化,並在所有修煉人的白細胞中都有變化的基因有 485個。235個基因的變化在6個修煉人呈不一致變化(有的上升有的下降,可能與個人修煉狀況有關)。有意思的是,其他250個基因呈同向變化,132個基因上調控,另外118個基因呈下調控。上升的基因多和免疫功 能有關(圖1),如抗微生物的小分子鈦-防禦素(Defensin-3, L12691)增加了6.7 倍 (圖3),干擾素及干擾素相關的、和受干擾素誘導的基因都明顯增加,而干擾素的抑制因子,Sarcolectin 卻明顯下降[48](圖2)。很明顯,調控趨勢是干擾素的功能增強。抗微生物的小分子在生物防禦系統中起著非常重要的作用,哺乳動物的防禦素是屬於體內表達量最高的廣譜抗菌素,在嗜中性白細胞中尤其高[49]。防禦素-3(A型防禦素)是一種帶負電荷的小分子鈦, 起著一種非氧化物性殺傷微生物的作用。A型防禦素通過裂解細菌的細胞膜起到一種先天免疫的作用[50,51]。最近,A型防禦素被認為是最重要的抗愛滋病毒的小分子(腳注)。干擾素-gamma,也是一種非常重要的抗病毒等微生物的細胞因子[52]。干擾素可通過誘導其他因子來調控免疫反應[53]。對轉基因小鼠和基因缺失小鼠的研究更進一步肯定了干擾素-gamma在免疫方面的作用[54,55]。這些基因的增高和功能方面的改變是相吻合的,嗜中性白細胞對細菌的吞噬和殺傷能力明顯增強 (圖4)。此外,I-309也明顯 增加(圖3)。 I-309 屬於趨化細胞因子,它的受體是愛滋病毒的輔助受體[56,57],因此,I-309 可能起著抗愛滋病毒感染的作用。I-309在修煉者體內普遍升高可能是修煉者對愛滋病毒有抵抗力的重要原因之一(根據一項未發表的文章)。當然,防禦素的增加可能是抗愛滋病毒感染的另外一個因素(腳注)。
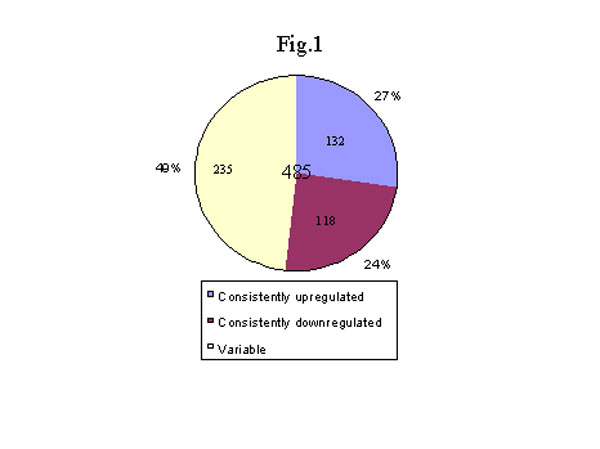
圖 1. 法輪功修煉者和正常對照組嗜中性白細胞基因表達變化總結。從亞裔法輪功修煉者(人數=6,平均年齡45.9歲,標準差14.3歲)和亞裔正常健康對照(人數=6,平均年齡40.3歲,標準差10.3歲)抽取30毫升血,肝素抗凝,操作方案由貝勒醫學院審查委員會通過。抽取靜脈血到肝素化的注射器,立刻分離細胞。用Ficoll-Hypaque密度梯度離心加右旋糖沉降法分離出多形核白細胞和單核細胞。為了避免進一步損傷,沒有除去紅細胞。將新鮮分離的嗜中性白細胞懸浮在磷酸緩衝液中,用Coulter細胞計數儀計數。顯微鏡檢測證實有96%到98%的細胞是嗜中性白細胞,其中95%的細胞可被苔盼藍排除法染色確認。用Qiagen試劑盒提取和純化總核糖核酸。5微克總核糖核酸, T7-d(T)24引物(GGCCAGTGAATTGTAATACGACTCACTATA GGGAGGCGG-(dT)24)和Superscript II (GIBCO-BRL, Rockvill, MD) 用於合成第一條cDNA鏈。第二條cDNA鏈的合成反應在16℃進行。反應體系中加入大腸桿菌DNA連接脢,大腸桿菌DNA多聚脢I,核糖核酸脢H,然後加入T4 DNA多聚脢以補齊新合成的cDNA鏈的末端。cDNA再經PLG膠-酚/氯仿抽提純化,乙醇沉澱。將純化了的cDNA在37℃離體轉錄反應中培養6小時以產生生物素標記的cRNA。將20微克cRNA放在緩衝液(含200毫摩爾Tris-Acetate, pH 8.1; 500毫摩爾醋酸鉀,150毫摩爾醋酸鎂)中,95C培養35分鐘使之成為片段。cRNA片段和經過預平衡的Affymetrix生物晶片在45℃雜交 14至16小時。雜交後移去反應液,用低強度緩衝液(6X標準磷酸鹽溶液加EDTA,0.01% Tween 20, 0.005%抗泡沫劑)洗10次,然後用高強度緩衝液(100 毫摩爾 MES, 0.1 摩爾氯化鈉,0.01% Tween 20)洗4次,用SAPE染色。用生物素標記的小鼠抗Avidin抗體孵育後,再用SAPE染色。生物晶片用惠普晶片掃描儀探測雜交信號。雜交信號生成的文本文件輸入到微軟Excel進行資料分析以找出法輪功修煉者和正常對照組之間變化達兩倍以上的信號。檢測了四個法輪功修煉者的12,000個基因。有 485個基因發生了變化,其中132個基因(佔變化數的27%)持續下調,118個基因(佔24%)持續上調。
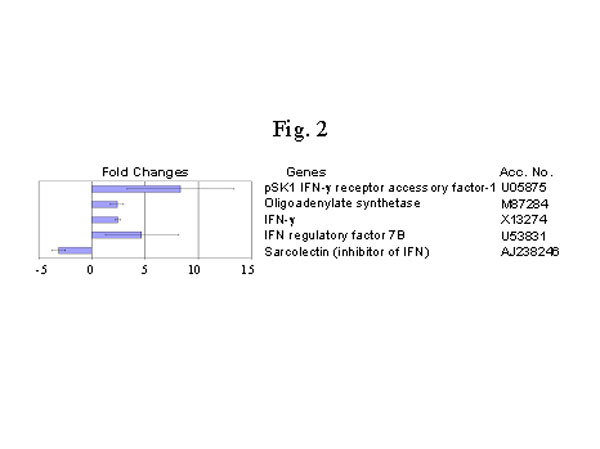
圖2. 干擾素相關基因的變化。四個法輪功修煉者和四個正常人干擾素相關基因變化的比較。數據是以mean + SD 表示。正數表示增加的倍數,負數表示減少的倍數。
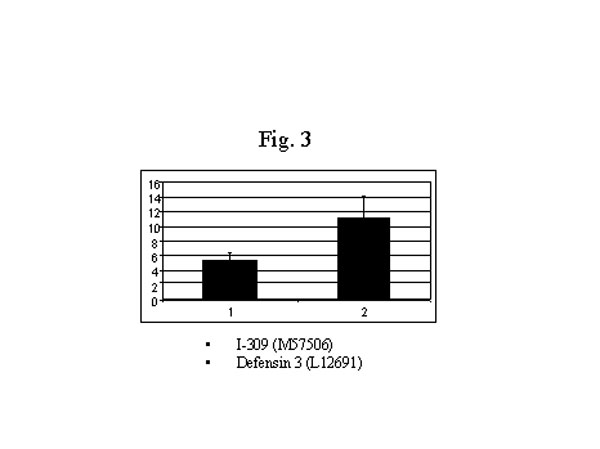
圖3. 抗微生物鈦基因的上調節。法輪功修煉者I-309和防禦素的變化。材料和方法見圖1說明。
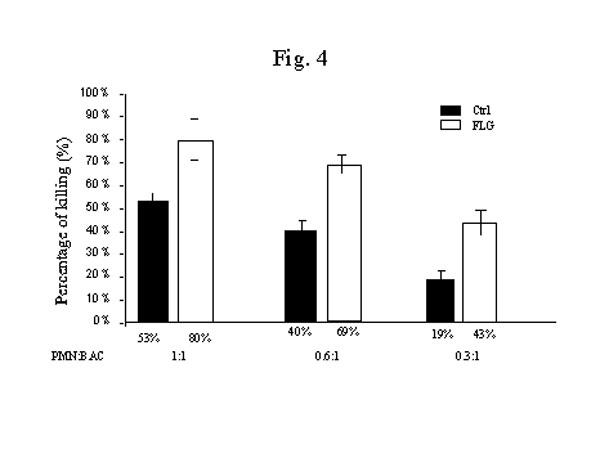
圖 4. 法輪功修煉者嗜中性白細胞的抗菌活性增加。葡萄球菌(SA)用作殺菌活性檢測的底物靶細胞。選用對數生長中期的葡萄球菌。將靶細胞葡萄球菌和嗜中性白細胞以不同的作用物/底物比混合,在37℃培養2個小時。2小時後作系列稀釋塗瓊脂盤,數菌落數以定量殺菌活性。用細菌和嗜中性白細胞混合培養後的活菌減少數來定量嗜中性白細胞的最大殺菌活性。細菌的最終濃度以瓊脂盤上的菌落數計算。殺菌活性以殺死細菌的百分比計。如圖所示,與正常對照組相比,法輪功組的嗜中性白細胞殺菌活性有顯著增加,殺傷力的增加在白細胞:細菌任何比例下都有顯著的統計學差異(P<0.0001)。
然而,免疫反應就像一柄雙刃劍。當它持續反應時,過度的免疫系統的激活就可能對主體有害[58]。那麼修煉人的免疫系統是如何解決這一難題的呢?我們發現了另一種超常的調控,BCl-2和BCl-XL,這些調控細胞死亡的基因表現為顯著地下調控(圖5)。 BCl-2和BCl-XL屬於BCl-2家族的抗細胞死亡成員[59,60]。這些抗細胞死亡基因的減少可能使修煉人的嗜中性白血球在受聚酯糖(lipopolysaccharide, LPS) 時加速死亡(圖6a和圖7)。LPS是一種炎症促進劑,能夠刺激細胞來表達細胞各種各樣的炎症因子[60]。很多慢性炎症損傷與受刺激的嗜中性白血球的死亡延遲有關[61]。而誘導細胞死亡卻有利於控制炎症 [62]。 另外,細胞死亡誘發的基因顯著地卻反向上調控,其中包括腫瘤壞死因子受體-1 相關蛋白(TNF-receptor-1 associatedprotein), BH3 相互作用基團死亡促進因子(BH3 interacting domain death agonist), 白細胞介素-1 轉換脢(Interleukin-1 convertase), 和白細胞介素-1轉換脢2 ( Interleukine-1-converting enzyme2)。這些促進細胞死亡的因子的增加可能是修煉人受刺激的細胞加速死亡的另一個因素(圖5)。
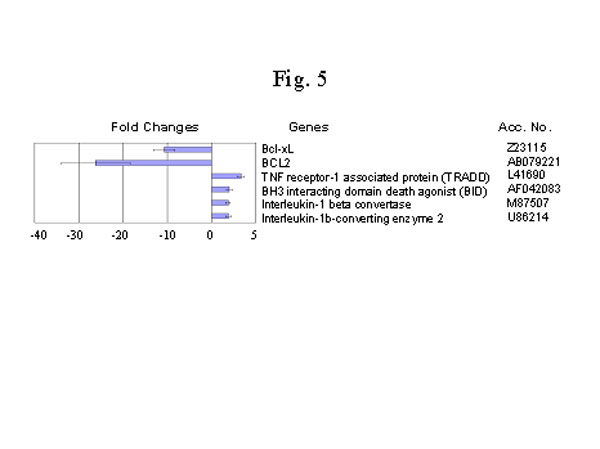
圖5. 細胞凋零死亡相關基因的變化。材料和方法見圖1說明。

圖 6. 法輪功修煉者嗜中性白細胞凋零死亡的變化。嗜中性白細胞以適當濃度重新混勻在含10%小牛血清的RPMI1640培養液中。吉姆薩染色證實分離出的多形核白細胞純度高於80%,其中的單核細胞少於1%。台盼藍排除法證實活細胞高於98%。刺激細胞用內毒素(25微克/毫升)。嗜中性白細胞在有或沒有內毒素刺激的條件下培養不同時間。細胞死亡用Roche公司的檢測盒測定。具體做法是,1x10^4 嗜中性白細胞培養在200微升含3%小牛血清的RPMI 1640培養液中,置於96孔培養板中37℃培養,在不同的時間點收樣。離心收集細胞,將細胞重新懸置在溶細胞液中,然後離心收集完整的細胞核沉澱。將含核小體的上清液移到Streptavidin包被的96孔脢聯板,與生物素標記的抗組蛋白抗體,超氧歧化脢標記的抗-DNA抗體共同孵育以形成抗體-核小體複合物。洗後,加入超氧歧化脢底物ABST孵育。在405納米光下測定吸收。這個脢鏈反應的結果顯示法輪功修煉者的細胞凋零死亡在內毒素刺激下有顯著增加而在沒有內毒素時顯著減少。* 表示均值有顯著性差異,P值小於0.01。t檢驗。資料來自7名法輪功修煉者和7名正常對照,採樣時間點是0,2,4,8和16小時。
細胞死亡有關基因的改變可能導致了煉功人嗜中性白細胞中表現的LPS刺激死亡的加速(圖7)。嗜中性白細胞的加速死亡能引起炎症的迅速消失,從而限制了由於免疫系統的增強而引起的自我損傷[63-65]。如圖6和圖7所示,在沒有LPS刺激的情況下,煉功人嗜中性白細胞的存活有顯著的延長,而這種細胞壽命的延長也可能與煉功所導致的新陳代謝水平下降有關。在非炎症的狀況下,決定嗜中性白細胞在非炎症情況下自發性死亡的因素是氧化自由基,而不是致死基因 [66,67]。我們的數據顯示法輪功及其它打坐修煉者的嗜中性白細胞受活性氧化劑的干預下降。有其他研究也說明血脂過氧化程度的下降可能跟打坐入定引起的緊張狀態降低密切相關[68]。
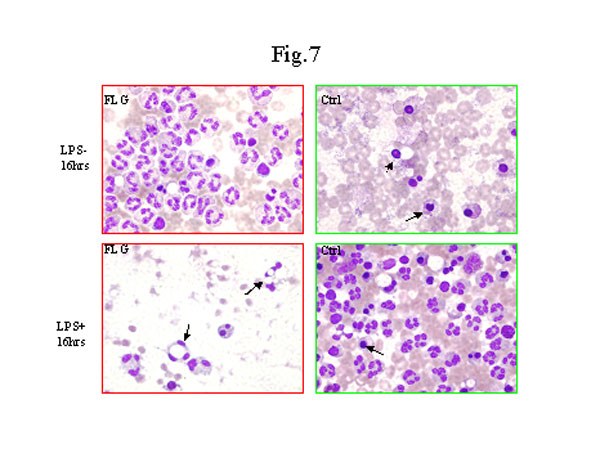
圖 7. 致炎因子內毒素對嗜中性白細胞凋零死亡的影響。分離和處理嗜中性白細胞的方法如圖6說明。不同時間點取樣的細胞經Weight法染色在光鏡下檢查,凋零死亡的細胞由紅圈圈出。箭頭所指的嗜中性白細胞表現出染色質濃縮,核變園,胞漿空泡形成和細胞縮小。嗜中性白細胞在有或沒有內毒素刺激的條件下培養16小時。如圖所示,法輪功修煉者的嗜中性白細胞在沒有內毒素刺激時大多數仍存活,受刺激時卻大部份出現了凋零死亡。與此相反,正常對照組的嗜中性白細胞在沒有內毒素刺激時是凋零死亡,受刺激時則存活。
免疫耐受和免疫激活是對立而又相互依賴的。免疫耐受可以保護宿主不被自我損傷,而免疫監視/激活則是消滅受損和變異的細胞並保護機體不被變異的細胞影響和病原體侵入。不過,陰中還必須有陽,因為只有免疫耐受而沒有免疫激活能導致機體易患癌症和感染性疾病;反之,陽中也必須有陰,因為沒有自我限制的免疫激活會導致嚴重的自我損傷。
法輪功修煉者的嗜中性白細胞表現出致炎基因(抗凋零死亡基因)的減少和炎症傾向得以限制(陰),同時免疫細胞的免疫力增強(陽)是為陽中有陰。與免疫力增強有關的改變代表了陰中陽的部份在細胞處於低活性時(低代謝)保護機體免於微生物侵犯。修煉者的嗜中性白細胞是低代謝低致炎的,卻具有免疫力增強的分子,是為陰中有陽。由於缺乏抗凋零死亡基因,激活的炎症細胞會被快速排除,由免疫力上調節導致的不利因素因而被抵消。
中醫認為,陰陽調和為健康之本。陰陽調和這一簡單卻極為重要的法則在健康、醫學和大自然中到處都能體現出來。陰陽理論也可用來解釋現代免疫學的概念,如免疫耐受性和免疫活性。免疫耐受(陰)和激活(陽)的平衡是一個生死攸關的問題。免疫耐受和激活是對立而又相互依賴(相生相剋)。免疫系統活性過低的話,會使免疫監督不能正常進行,從而造成突變和炎症細胞不能及時排除,這將導致癌症和慢性炎症等。但免疫監督/激活(陽)又不能過強,因為那會造成對自身的傷害,所以就需要自我限制對其的制約。比如,炎症細胞必須用細胞死亡的調控來清除,從而消除和停止炎症反應(陽中有陰)。免疫耐受在使生命保持和諧性方面非常重要,但僅僅耐受而沒有免疫監督/激活化會導致感染,癌症,或自身免疫疾病,這是陰陽相互作用的典型表現。因此,陽中必須有陰。由身心相互作用引起的調控是一種陰陽調和的平衡,從而使得免疫系統的「雙刃劍」的特性得到糾正,也就是說免疫力可以上升而不會有反作用。在精神層面上修煉的人,他們嗜中性白細胞中致死基因的改變支持了這種假說。氣功練習者嗜中性白細胞中抗細胞死亡基因的下調控促成了炎症細胞的迅速清除,而健康嗜中性白細胞的存活期的延長,則可能是由於打坐入定引起的細胞代謝水平下降從而使氧化自由基的減少所致,這種雙向調控有效地保持了免疫的陰陽平衡。打坐入定所造成的代謝水平下降(見下文)而使機體處於一種低反應狀態(陰),免疫活性就會增強(陽),干擾素和防禦素增加及吞噬功能增強)從而來平衡保護這種低反應狀態。這種雙向調控的現象將會為我們治療方針另闢蹊徑,如果免疫反應能夠下調而又不影響整體免疫力,就能對移植排斥,自身免疫疾病和過敏等的治療有極大的幫助[69]。反之,如果基於陰陽調和基礎上的免疫調控可能會是治療多種免疫疾病的又一種選擇。
新陳代謝率的降低和細胞「垃圾箱」的減少
細胞的新陳代謝高低決定了蛋白質合成和降解的速率。然而,蛋白質合成和降解的經濟性長期以來一直被人忽視,直到重新理解泛素依賴性的蛋白質降解系統 (Ubiquitin-dependent protein degradation pathway, UPP)[70]。在真核細胞中損壞的,異化的,和老化的蛋白質的主要處理系統是UPP[71]。在正常人的細胞中,大量新合成的不正常的蛋白質都會通過 UPP來降解,這可能是因為正常人細胞的新陳代謝率也增高所致。例如,一個細胞每分鐘能合成2E6個核糖體蛋白質的拷貝,其中約有超過30%新合成的蛋白質立即就被UPP銷毀了。在病理條件下,被送入UPP降解的蛋白質會顯著增加,大量的人類疾病可能和UPP都有關係[71-74]。
UPP 可以通過修飾細胞內各種各樣的蛋白質,參與很多重要的,基本的細胞活動。這其中包括細胞周期,分裂,分化,和發生發育的調控,細胞對刺激和外界因子的反應的調控,神經系統的形態發生,細胞表面的受體和離子通道的下調控,蛋白質分泌的定量調控,DNA修復,免疫和炎症反應的調控,以及細胞器的生成等等。已確定的要通過UPP的細胞蛋白質有幾百之多,而且這個名單還在穩步增長。蛋白質基質的多樣性和結構的複雜性,使得這一系統和許多重要疾病的發病機理聯繫起來,包括基因疾病、神經細胞退行性疾病、癌症、肌萎縮、高血壓、白血病、自身免疫疾病、炎症、老年性失調。最近,有初步試驗結果表明, UPP的蛋白脢的抑制劑可能對癌症有治療作用[75,76]。很多情況下,與UPP有關的疾病都是因為過高的新陳代謝率。它就像一個分子傳感器,其活性會隨著外界和內在的應力增加而變化。因為這個體系就像垃圾處理站,總是把反常的和破損的細胞拖進來分解掉,我們把它比喻成一個細胞中的黑洞。
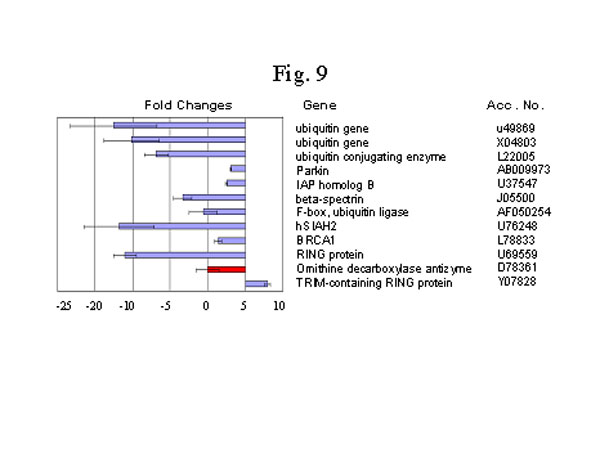
圖9. UPP調節基因的下調節。材料和方法見圖1說明。
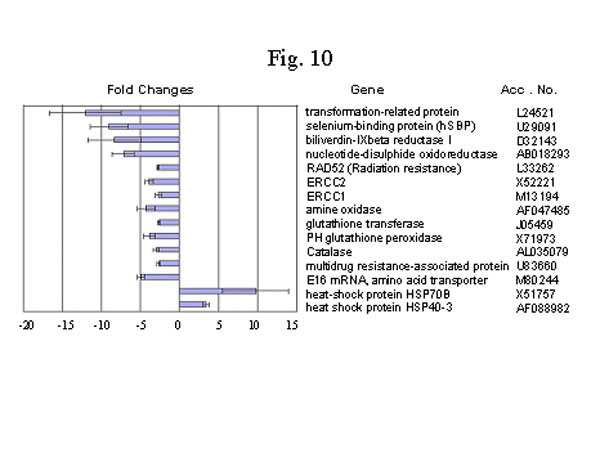
圖10. 緊張相關基因的改變。材料和方法見圖1說明。

圖11. 核小體蛋白相關基因的下調節。材料和方法見圖1說明。
UPP 的主要成份是泛素,UPP系統有三類主要的脢:E1(ubiquitin activating enzyme, 泛素激活脢),E2(ubiquitin conjugating enzyme, 泛素交聯脢)和E3(ubiquitin ligase, 泛素連接脢)70。RING(Really Interesting New Gene, 誘人的新基因)RING-finger蛋白質是一組近來才明確其特殊功能的蛋白質[77]。越來越多的研究表明在特定的泛素反應中RING-finger 蛋白質起著主導作用,而且可能所有的RING-finger蛋白質都是E3型泛素-蛋白聯結脢[78,79]。我們發現大法修煉者身上分離出的嗜中性白細胞,其泛素、E2和E3的基因表達量有明顯的下調控趨勢(圖9)。在所有我們確認的RING-finger蛋白質中,只有屬於RBCC家族的含TRIM 的RING 蛋白質是上調控的[80]。相當有意思的是,有7個泛素E3基因同時下調(圖9)。此外,一種與UPP無關的蛋白質降解脢,ornithine decarboxylase antizyme [81],也有顯著減少(圖10)。UPP系統的下調可能是因新陳代謝率的下降而非打坐入定的直接效應。據報導,泛素是一個緊張或熱休克調控的基因[82,83]。不過,目前沒有文獻報導UPP系統中其它的成分也能隨緊張或熱休克調控。以下描述的其它與緊張刺激有關的基因的變化也說明UPP體系的下調控可能是個非直接過程。
其它下調控的基因包括,參與蛋白質合成的基因,DNA修補的抗氧化脢。主要的抗氧化脢,過氧化氫脢和谷胱甘鈦過氧化氫脢,也減少了(圖10)。這些與緊張刺激相關的關鍵脢的減少,以及其它受緊張刺激調控的基因下降,意味著氧化物的生成和大分子的破壞減少,這些基因包括酵母基因在哺乳動物體內的同源基因RAD52(抗輻射)和ERCCs(切除修補)。核糖體是蛋白質合成中重要的成分,而11個核糖體基因中10個下降說明蛋白質合成也可能下降(圖11)。令人意想不到的是,兩個熱休克蛋白質,HSP70和HSP40-3,卻顯著上升, 這與其它對緊張刺激有關的基因的下降恰恰相反(圖10)。 這些緊張刺激調控基因反向調控的分子機理還需要進一步確定。但對於類似這些熱休克基因,研究是否可以通過上調控來起到保護作用是應該很有意思的。有證據表明這些熱休克蛋白可能對心臟確有保護作用[84]。
這一項用DNA微排列技術研究在基因的水平上研究基因表達狀況說明,煉功人細胞的代謝率有下降趨勢。UPP系統的下調可以用兩種理由解釋,1)底物依賴性的下調,因為代謝率的下降引起代謝產物的減少導致了廢物處理UPP過程的下調,2)打坐入定引發的細胞活動某種程度上直接導致了UPP基因表達的UPP系統的某些分子轉錄水平下降。對於UPP系統的下調可能與修煉人的健康表型有關這一問題,目前還沒有文獻報導予以支持,但某些在嚙齒類及靈長類動物身上進行的限制熱量攝入實驗的結果可能支持這一發現。限制熱量攝入(Caloric restriction, CR)是已知的唯一能夠使嚙齒類乃至靈長類動物減緩衰老和延長壽命的方法[85,86]。CR小鼠的壽命延長的表型也可能與新陳代謝的下降及UPP活性的下調有關[87,88]。凡此種種都支持新陳代謝的下降也能相應地導致「垃圾處理器」的減小這樣一種觀點。此外,有的科學家還發現CR動物具有抗應力和疾病抵抗能力增強這一表型[89,90]。但是否氣功修練者的抵抗力增強與此有關還有待進一步證實。修煉者的泛素蛋白質降解系統的劇烈變化說明此體系可能作為一個新陳代謝變化的分子感應器。鑑於泛素蛋白質降解系統在各種細胞生理和病理過程中的重要性,法輪大法修煉或許可作為針對多種人類疾病的替代療法。
最後,我們小結一下以上的發現。精神修煉可能影響下列基因的表達,其中包括蛋白質合成與降解,細胞代謝,細胞死亡,以及免疫調控。此外,我們的結果也說明周圍血細胞可以被用來研究基因水平上的基因表達變化。在這些變化中,最值得一提的是細胞凋零死亡基因的調控:抗細胞凋零死亡基因的下調控有利於免疫增強的情況下炎症細胞的快速解決。這一調控非常重要,因為可以避免由於免疫力增強而造成對個體的自我損傷。與代謝率下降有關的趨勢支持長期以來的看法,即降低代謝率可能減緩老化過程和延長壽命。UPP系統的顯著變化進一步說明了這一系統在生理和病理方面的重要意義,也反應了這一系統在人類精神意識方面對健康的重要性。目前,為了進一步研究人類的精神意識如何影響基因表達,我們正在進行修煉前後的對照,以及其他精神修煉的變化。我們希望我們的研究為現代身心相互作用方面提供一些新的研究策略和方向。
參考文獻
1. Danner DD, Snowdon DA, Friesen WV. Positive emotions in early life and longevity: findings from the nun study. J Pers Soc Psychol 2001;80(5):804-13.
2. Kiecolt-Glaser JK, McGuire L, Robles TF, Glaser R. EMOTIONS, MORBIDITY, AND MORTALITY: New Perspectives from Psychoneuroimmunology. Annu Rev Psychol 2002;53:83-107.
3. Editorial. Progress and promise in research on social and cultural demensions of health: a research agenda. In: NIH, editor; 2001; NIH, Bethesda, MD 20892.
4. King MC, Wilson AC. Evolution at two levels in humans and chimpanzees. Science 1975;188(4184):107-16.
5. Paabo S. Genomics and society. The human genome and our view of ourselves. Science 2001;291(5507):1219-20.
6. Enard W, Khaitovich P, Klose J, Zollner S, Heissig F, Giavalisco P, Nieself-Struwe K, Muchmore E, Varki A, Ravid R and others. Intra- and interspecific variation in primate gene expression patterns. Science 2002;296:340-343.
7. Beecher H. The powerful placebo. JAMA 1955;159:1602-1606.
8. Shapiro AK. Placebo effects in medicine,psychotherapy, and psychoanalysis. In: Bergin A, Garfield S, editors. Handbook of psychotherapy and beharior change. New York: Wiley and Sons; 1971.
9. Benson H, Epstein MD. The placebo effect. A neglected asset in the care of patients. Jama 1975;232(12):1225-7.
10. Stefano GB, Fricchione GL, Slingsby BT, Benson H. The placebo effect and relaxation response: neural processes and their coupling to constitutive nitric oxide. Brain Res Brain Res Rev 2001;35(1):1-19.
11. Walsh BT, Seidman SN, Sysko R, Gould M. Placebo response in studies of major depression: variable, substantial, and growing. Jama 2002;287(14):1840-7.
12. Schweiger A, Parducci A. Nocebo: the psychologic induction of pain. Pavlov J Biol Sci 1981;16(3):140-3.
13. Spiegel H. Nocebo: the power of suggestibility. Prev Med 1997;26(5 Pt 1):616-21.
14. Benson H. The nocebo effect: history and physiology. Prev Med 1997;26(5 Pt 1):612-5.
15. Barsky AJ, Saintfort R, Rogers MP, Borus JF. Nonspecific medication side effects and the nocebo phenomenon. Jama 2002;287(5):622-7.
16. Hrobjartsson A, Gotzsche PC. Is the placebo powerless? An analysis of clinical trials comparing placebo with no treatment. N Engl J Med 2001;344(21):1594-602.
17. Petrovic P, Kalso E, Petersson KM, Ingvar M. Placebo and Opioid Analgesia--Imaging a Shared Neuronal Network. Science 2002;7:7.
18. de la Fuente-Fernandez R, Ruth TJ, Sossi V, Schulzer M, Calne DB, Stoessl AJ. Expectation and dopamine release: mechanism of the placebo effect in Parkinson's disease. Science 2001;293(5532):1164-6.
19. Leuchter AF, Cook IA, Witte EA, Morgan M, Abrams M. Changes in brain function of depressed subjects during treatment with placebo. Am J Psychiatry 2002;159(1):122-9.
20. Vedantam S. Against depression, a sugar pill is hard to beat: Placebos improve mood, change brain chemistry in majority of trials of antidepressants. Washington post 2002.
21. Torrey EF, Miller J. The invisible plague: The rise of mental illness from 1750 to the present; 2001.
22. Schuster MA, Stein BD, Jaycox L, Collins RL, Marshall GN, Elliott MN, Zhou AJ, Kanouse DE, Morrison JL, Berry SH. A national survey of stress reactions after the September 11, 2001, terrorist attacks. N Engl J Med 2001;345(20):1507-12.
23. Galea S, Ahern J, Resnick H, Kilpatrick D, Bucuvalas M, Gold J, Vlahov D. Psychological sequelae of the September 11 terrorist attacks in New York City. N Engl J Med 2002;346(13):982-7.
24. Vlahov D, Galea S, Resnick H, Ahern J, Boscarino JA, Bucuvalas M, Gold J, Kilpatrick D. Increased use of cigarettes, alcohol, and marijuana among Manhattan, New York, residents after the September 11th terrorist attacks. Am J Epidemiol 2002;155(11):988-96.
25. Goodnouch A. Post-9/11 pain found to linger in young minds. New York Times 2002 May 20.
26. AP. Post-trauma stress plagues NYC kids. MSNBC 2002;https://www.msnbc.com/news/746411.asp.
27. Jacobs GD. The physiology of mind-body interactions: the stress response and the relaxation response. J Altern Complement Med 2001;7(Suppl 1):S83-92.
28. Tabor HK, Risch NJ, Myers RM. OPINION: Candidate-gene approaches for studying complex genetic traits: practical considerations. Nat Rev Genet 2002;3(5):391-7.
29. Kiberstis P, Roberts L. It's Not Just the Genes. Science 2002;296(5568):685-.
30. Marx J. Unraveling the causes of diabetes. Science 2002;296(5568):686-9.
31. Marshall E. Lupus: Mysterious Disease Holds Its Secrets Tight. Science 2002;296(5568):689-691.
32. Sawa A, Snyder SH. Schizophrenia: diverse approaches to a complex disease. Science 2002;296(5568):692-5.
33. Willett WC. Balancing Life-Style and Genomics Research for Disease Prevention. Science 2002;296(5568):695-698.
34. Rees J. Complex Disease and the New Clinical Sciences. Science 2002;296(5568):698-700.
35. Strohman R. Maneuvering in the Complex Path from Genotype to Phenotype. Science 2002;296(5568):701-703.
36. Editorial. Prevention remains the best medicine. Nature 2002;416(6880):416.
37. Snowdon DA, Greiner LH, Markesbery WR. Linguistic ability in early life and the neuropathology of Alzheimer's disease and cerebrovascular disease. Findings from the Nun Study. Ann N Y Acad Sci 2000;903:34-8.
38. Snowdon DA. Aging with grace: what the nun study teaches us about leading longer, healthier, and more meaningful lives. New York, New York: Bantam Books; 2001.
39. Gimbel MA. Yoga, meditation, and imagery: clinical applications. Nurse Pract Forum 1998;9(4):243-55.
40. Herrick CM, Ainsworth AD. Invest in yourself. Yoga as a self-care strategy. Nurs Forum 2000;35(2):32-6.
41. Pettinati PM. Meditation, yoga, and guided imagery. Nurs Clin North Am 2001;36(1):47-56.
42. Lewis DE. T'ai chi ch'uan. Complement Ther Nurs Midwifery 2000;6(4):204-6.
43. Chen KM, Snyder M, Krichbaum K. Clinical use of tai chi in elderly populations. Geriatr Nurs 2001;22(4):198-200.
44. Li HZ. Zhuan Falun - The Complete Teaching of Falun Gong. Gloucester: Fair Winds Press; 2001. 345 p.
45. Coker KH. Meditation and prostate cancer: integrating a mind/body intervention with traditional therapies. Semin Urol Oncol 1999;17(2):111-8.
46. Shang C. Emerging paradigms in mind-body medicine. J Altern Complement Med 2001;7(1):83-91.
47. Jones BM. Changes in cytokine production in healthy subjects practicing Guolin Qigong : a pilot study. BMC Complement Altern Med 2001;1(1):8.
48. Kaba A, Jiang PH, Chany-Fournier F, Chany C. Sarcolectin (SCL): structure and expression of the recombinant molecule. Biochimie 1999;81(7):709-15.
49. Kaiser V, Diamond G. Expression of mammalian defensin genes. J Leukoc Biol 2000;68(6):779-84.
50. van Wetering S, Sterk PJ, Rabe KF, Hiemstra PS. Defensins: key players or bystanders in infection, injury, and repair in the lung? J Allergy Clin Immunol 1999;104(6):1131-8.
51. Fellermann K, Stange EF. Defensins -- innate immunity at the epithelial frontier. Eur J Gastroenterol Hepatol 2001;13(7):771-6.
52. Samuel CE. Antiviral actions of interferons. Clin Microbiol Rev 2001;14(4):778-809.
53. Shtrichman R, Samuel CE. The role of gamma interferon in antimicrobial immunity. Curr Opin Microbiol 2001;4(3):251-9.
54. Jouanguy E, Doffinger R, Dupuis S, Pallier A, Altare F, Casanova JL. IL-12 and IFN-gamma in host defense against mycobacteria and salmonella in mice and men. Curr Opin Immunol 1999;11(3):346-51.
55. Dorman SE, Holland SM. Interferon-gamma and interleukin-12 pathway defects and human disease. Cytokine Growth Factor Rev 2000;11(4):321-33.
56. Horuk R, Hesselgesser J, Zhou Y, Faulds D, Halks-Miller M, Harvey S, Taub D, Samson M, Parmentier M, Rucker J and others. The CC chemokine I-309 inhibits CCR8-dependent infection by diverse HIV-1 strains. J Biol Chem 1998;273(1):386-91.
57. Lee S, Tiffany HL, King L, Murphy PM, Golding H, Zaitseva MB. CCR8 on human thymocytes functions as a human immunodeficiency virus type 1 coreceptor. J Virol 2000;74(15):6946-52.
58. Matzinger P. An innate sense of danger. Ann N Y Acad Sci 2002;961:341-2.
59. Adams JM, Cory S. Life-or-death decisions by the Bcl-2 protein family. Trends Biochem Sci 2001;26(1):61-6.
60. Smith JA. Neutrophils, host defense, and inflammation: a double-edged sword. J Leukoc Biol 1994;56(6):672-86.
61. Webb PR, Wang KQ, Scheel-Toellner D, Pongracz J, Salm
on M, Lord JM. Regulation of neutrophil apoptosis: a role for protein kinase C and phosphatidylinositol-3-kinase. Apoptosis 2000;5(5):451-8.
62. Ohta K, Yamashita N, Tajima M, Miyasaka T, Kawashima R, Nakano J, Arioka H, Ishii A, Horiuchi T, Miyamoto T. In vivo effects of apoptosis in asthma examined by a murine model. Int Arch Allergy Immunol 2001;124(1-3):259-61.
63. Simpson SJ, Hines PJ. Self-Discrimination, a Life and Death Issue. Science 2002;296(5566):297-.
64. Matzinger P. The Danger Model: A Renewed Sense of Self. Science 2002;296(5566):301-305.
65. Medzhitov R, Janeway CA, Jr. Decoding the patterns of self and nonself by the innate immune system. Science 2002;296(5566):298-300.
66. Akgul C, Moulding DA, Edwards SW. Molecular control of neutrophil apoptosis. FEBS Lett 2001;487(3):318-22.
67. Kasahara Y, Iwai K, Yachie A, Ohta K, Konno A, Seki H, Miyawaki T, Taniguchi N. Involvement of reactive oxygen intermediates in spontaneous and CD95 (Fas/APO-1)-mediated apoptosis of neutrophils. Blood 1997;89(5):1748-53.
68. Schneider RH, Nidich SI, Salerno JW, Sharma HM, Robinson CE, Nidich RJ, Alexander CN. Lower lipid peroxide levels in practitioners of the Transcendental Meditation program. Psychosom Med 1998;60(1):38-41.
69. Check E. Immunology: The virtue of tolerance. Nature 2002;418(6896):364-6.
70. Glickman MH, Ciechanover A. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev 2002;82(2):373-428.
71. Yewdell JW. Not such a dismal science: the economics of protein synthesis, folding, degradation and antigen processing. Trends Cell Biol 2001;11(7):294-7.
72. Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem 1998;67:425-79.
73. Vu PK, Sakamoto KM. Ubiquitin-mediated proteolysis and human disease. Mol Genet Metab 2000;71(1-2):261-6.
74. Ciechanover A, Schwartz AL. Ubiquitin-mediated degradation of cellular proteins in health and disease. Hepatology 2002;35(1):3-6.
75. Adams J. Proteasome inhibition in cancer: development of PS-341. Semin Oncol 2001;28(6):613-9.
76. Garber K. taking garbage in, tossing cancer out? Science 2002;295(5555):612-3.
77. Joazeiro CA, Weissman AM. RING finger proteins: mediators of ubiquitin ligase activity. Cell 2000;102(5):549-52.
78. Freeman H. An increase in insanity. Nature 2002;417(6885):123.
79. Jackson PK, Eldridge AG, Freed E, Furstenthal L, Hsu JY, Kaiser BK, Reimann JD. The lore of the RINGs: substrate recognition and catalysis by ubiquitin ligases. Trends Cell Biol 2000;10(10):429-39.
80. Reymond A, Meroni G, Fantozzi A, Merla G, Cairo S, Luzi L, Riganelli D, Zanaria E, Messali S, Cainarca S and others. The tripartite motif family identifies cell compartments. Embo J 2001;20(9):2140-51.
81. Coffino P. Antizyme, a mediator of ubiquitin-independent proteasomal degradation. Biochimie 2001;83(3-4):319-23.
82. Bond U, Schlesinger MJ. Ubiquitin is a heat shock protein in chicken embryo fibroblasts. Mol Cell Biol 1985;5(5):949-56.
83. Bond U, Schlesinger MJ. The chicken ubiquitin gene contains a heat shock promoter and expresses an unstable mRNA in heat-shocked cells. Mol Cell Biol 1986;6(12):4602-10.
84. Latchman DS. Heat shock proteins and cardiac protection. Cardiovasc Res 2001;51(4):637-46.
85. Lane MA, Black A, Handy A, Tilmont EM, Ingram DK, Roth GS. Caloric restriction in primates. Ann N Y Acad Sci 2001;928:287-95.
86. Roth GS, Ingram DK, Lane MA. Caloric restriction in primates and relevance to humans. Ann N Y Acad Sci 2001;928:305-15.
87. Lee CK, Klopp RG, Weindruch R, Prolla TA. Gene expression profile of aging and its retardation by caloric restriction. Science 1999;285(5432):1390-3.
88. Lee CK, Weindruch R, Prolla TA. Gene-expression profile of the ageing brain in mice. Nat Genet 2000;25(3):294-7.
89. Raffoul JJ, Guo Z, Soofi A, Heydari AR. Caloric restriction and genomic stability. J Nutr Health Aging 1999;3(2):102-10.
90. Yu BP, Chung HY. Stress resistance by caloric restriction for longevity. Ann N Y Acad Sci 2001;928:39-47.
腳 註﹕
文稿寄出後,又出現了新的證據提示防禦素可能也參與了煉功人的抗HIV作用。最近,張等人發現CD8的抗病毒因子就是防禦素(1,2,3),具有很強的抗 HIV-1的活性。(Zhang et al. Contribution of human alpha-defensin 1, 2, and 3 to the anti-HIV-1 activity of CD8 antiviral factor Science 2002 Nov 1;298(5595):995-1000)
致謝:
感謝弗傑尼亞Mason研究中心王同文的有益的建議;感謝貝勒醫學院的C.Wayne Smith討論,鼓勵和支持;感謝L.Schmidt和C.Robinson的文秘工作。感謝吳為的翻譯工作。這項研究的經費來自貝勒醫學院醫學系的系基金(封莉莉)。